IgG型治療性單抗(mAbs)的可結(jié)晶片段(Fc)區(qū)域常通過(guò)工程改造優(yōu)化其藥理學(xué)和藥代動(dòng)力學(xué)(PK)特性,但也可能對(duì)安全性產(chǎn)生影響�����。
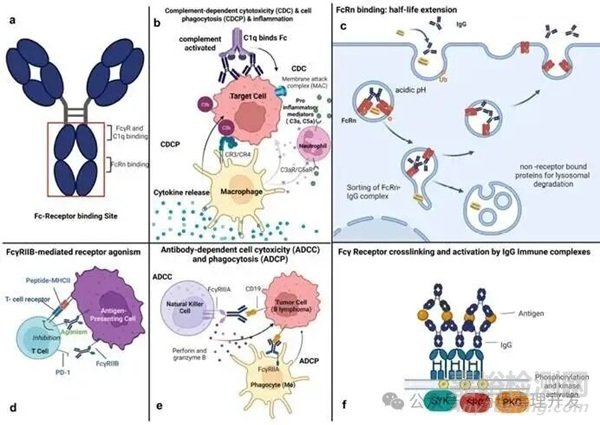
通過(guò)增強(qiáng)與激活型Fcγ受體(FcγRs)或補(bǔ)體成分1q(C1q)的結(jié)合能力����,可實(shí)現(xiàn)特定效應(yīng)功能的增強(qiáng)。Fc工程改造的IgG單抗能增強(qiáng)對(duì)靶細(xì)胞的抗體依賴性細(xì)胞毒性(ADCC)�、抗體依賴性細(xì)胞吞噬作用(ADCP)、補(bǔ)體依賴性細(xì)胞毒性(CDC)或補(bǔ)體依賴性細(xì)胞介導(dǎo)的細(xì)胞毒性(CDCC)�����。此外�����,通過(guò)氨基酸取代促進(jìn)Fc-Fc六聚化和FcγRIIB相互作用,可實(shí)現(xiàn)基于受體聚集的激動(dòng)作用���。
除了增強(qiáng)����,F(xiàn)c改造還有很多其它方向:1)Fc段沉默:通過(guò)降低FcγR和C1q的結(jié)合能力����,使Fc結(jié)構(gòu)域(部分)沉默;2)Fc段多聚化:將Fc結(jié)構(gòu)域多聚化��,以阻斷FcγRs和新生兒Fc受體(FcRn)�;3)調(diào)控半衰期:改造Fc結(jié)構(gòu)域以增強(qiáng)或減弱其與FcRn的結(jié)合�����,從而精細(xì)調(diào)節(jié)IgG的半衰期����。
由于工程改造策略的復(fù)雜性,且經(jīng)改造的單抗在最常用的藥理毒理學(xué)種屬(即食蟹猴和嚙齒類(lèi)動(dòng)物)中受體結(jié)合親和力發(fā)生改變���,因此對(duì)這類(lèi)改造單抗進(jìn)行充分的非臨床藥理學(xué)和安全性評(píng)估的需求日益增加��。
本文介紹了人類(lèi)與非臨床常見(jiàn)種屬中的FcR表達(dá)及功能����。
人類(lèi)FcR表達(dá)及功能
人類(lèi)FcγR的表達(dá)及功能
從功能上看,人類(lèi)FcγR可分為激活型和抑制型受體����,它們分別通過(guò)細(xì)胞內(nèi)基于免疫受體酪氨酸的激活基序(ITAMs)或抑制基序(ITIMs)傳遞信號(hào),在與結(jié)合靶標(biāo)的IgG抗體交叉結(jié)合后影響細(xì)胞功能��。
人類(lèi)FcγR的分類(lèi)與結(jié)合特性
激活型受體:人類(lèi)hFcγRI(CD64)、hFcγRIIA(CD32a)�、hFcγRIIC(CD32c)和hFcγRIIIA(CD16a)均為激活型受體。
抑制型受體:FcγRIIB(CD32b)是唯一的抑制型受體����。
結(jié)合親和力:大多數(shù)FcγR與IgG1 Fc的親和力較低,但當(dāng)多個(gè)抗體聚集形成免疫復(fù)合物時(shí)���,可形成強(qiáng)烈的功能性相互作用����。這種聚集使FcγR發(fā)生交聯(lián),這是觸發(fā)下游免疫信號(hào)和功能的關(guān)鍵��。高親和力受體FcγRI屬于例外情況���,它可結(jié)合單體和非聚集型IgG��。
特殊類(lèi)型:FcγRIIIB(CD16b)較為特殊�,它缺乏跨膜和胞質(zhì)部分,因此無(wú)直接信號(hào)活性�,但可能通過(guò)糖基磷脂酰肌醇(GPI)錨定在細(xì)胞膜上。由于其在大量中性粒細(xì)胞上的高表達(dá)����、與FcγRIIIA的序列相似性,以及在小鼠和大多數(shù)非人靈長(zhǎng)類(lèi)動(dòng)物(NHP)中不表達(dá)的特點(diǎn)����,需對(duì)其潛在功能進(jìn)行審慎考量���。
人類(lèi)FcγR的表達(dá)分布與動(dòng)態(tài)變化
表達(dá)范圍:FcγRs在人類(lèi)所有免疫細(xì)胞亞群和血小板上均有表達(dá)��。
表達(dá)差異:部分細(xì)胞僅表達(dá)一種受體����,因此傾向于以極化方式發(fā)揮作用(例如,人類(lèi)B細(xì)胞僅表達(dá)抑制型FcγRIIB�;大多數(shù)人類(lèi)NK細(xì)胞僅表達(dá)激活型
FcγRIIIA)。另一些細(xì)胞則表達(dá)多種受體��,因此免疫復(fù)合物的大小和同種型會(huì)導(dǎo)致激活型與抑制型受體信號(hào)的動(dòng)態(tài)競(jìng)爭(zhēng)���。例如���,人類(lèi)巨噬細(xì)胞和樹(shù)突狀細(xì)胞以各種組合方式表達(dá)激活型和抑制型FcγR。
動(dòng)態(tài)調(diào)節(jié):FcγR的表達(dá)具有動(dòng)態(tài)性��,可被IFN-γ�����、TNF����、IL-4���、IL-10或IL-13等細(xì)胞因子上調(diào)。例如���,IFN-γ可強(qiáng)效且差異性地誘導(dǎo)未成熟DCs上激活型FcγRIIa與抑制型FcγRIIb的表達(dá)���;部分FcγR在與免疫復(fù)合物(ICs)相互作用后還會(huì)被內(nèi)化。
IgG的關(guān)鍵效應(yīng)功能
NK細(xì)胞上的FcγRIIIA可促進(jìn)ADCC介導(dǎo)的靶細(xì)胞殺傷�。
巨噬細(xì)胞上的FcγRIIA和FcγRIIIA以及中性粒細(xì)胞上的FcγRIIIB可促進(jìn)ADCP及后續(xù)對(duì)抗體結(jié)合靶標(biāo)的殺傷。
FcγR(如FcγRIIB)的結(jié)合還可通過(guò)促進(jìn)IgG的Fab段與受體的交聯(lián)/激動(dòng)作用�����,增強(qiáng)靶效應(yīng)細(xì)胞的激活����。一個(gè)細(xì)胞上的FcγRIIB對(duì)IgG的“超交聯(lián)”可在靶細(xì)胞中產(chǎn)生信號(hào)(如凋亡或激活)。IgG與FcγRIIB的交聯(lián)可抑制B細(xì)胞激活����。吞噬細(xì)胞和肝血竇內(nèi)皮細(xì)胞可通過(guò)FcγRIIB參與小免疫復(fù)合物的“清除”或內(nèi)化。
血小板或中性粒細(xì)胞上的FcγRIIA可導(dǎo)致可溶性介質(zhì)的分泌����、凝血或中性粒細(xì)胞胞外陷阱形成。
FcγRIIIB被認(rèn)為在清除循環(huán)免疫復(fù)合物中發(fā)揮作用����,且不引起中性粒細(xì)胞激活。但在某些條件下���,hFcγRIIIB可與hFcγRIIA等其他信號(hào)分子協(xié)同作用��,參與免疫復(fù)合物介導(dǎo)的中性粒細(xì)胞激活����。
補(bǔ)體系統(tǒng)的作用
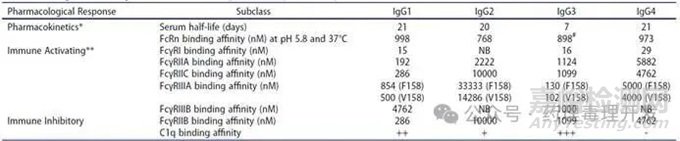
補(bǔ)體系統(tǒng)通過(guò)經(jīng)典途徑��、凝集素途徑或替代途徑“補(bǔ)充”體液免疫和細(xì)胞免疫功能���,包括直接防御病原體�、促進(jìn)炎癥反應(yīng)和清除免疫復(fù)合物����。IgG可結(jié)合C1q以激活經(jīng)典補(bǔ)體途徑并驅(qū)動(dòng)CDC,不同IgG亞類(lèi)的活性存在差異(IgG1和IgG3>IgG4)����。補(bǔ)體激活還會(huì)產(chǎn)生C3b以及過(guò)敏毒素C3a和C5a���,這些分子可通過(guò)結(jié)合巨噬細(xì)胞和中性粒細(xì)胞上的C3bR、C3aR和C5aR����,使靶細(xì)胞被調(diào)理吞噬����。
非臨床動(dòng)種屬中的FcγR表達(dá)及功能
在非臨床評(píng)估中,必須考慮動(dòng)物與人類(lèi)在FcγR表達(dá)及功能上的差異所帶來(lái)的轉(zhuǎn)化影響�。
非人靈長(zhǎng)類(lèi)動(dòng)物
食蟹猴和恒河猴的FcγRI、FcγRIIIA和FcγRIIA在表達(dá)����、功能及與人類(lèi)、食蟹猴和恒河猴IgG1-IgG4的結(jié)合模式上��,均表現(xiàn)出一定相似性���。例如����,食蟹猴血小板上表達(dá)FcγRIIA,因此通?���?稍诤镒由砩显u(píng)估通過(guò)FcγRIIA結(jié)合引發(fā)的人類(lèi)血小板激活風(fēng)險(xiǎn)����。但二者也存在一些差異,比如:1)獼猴體內(nèi)不存在FcγRIIC�����;2)獼猴粒細(xì)胞不表達(dá)FcγRIIIB�,但FcγRII的表達(dá)水平高于人類(lèi)細(xì)胞;3)人類(lèi)IgG1和IgG3與食蟹猴及人類(lèi)FcγRs的結(jié)合能力強(qiáng)于人類(lèi)IgG2和IgG4�,但人類(lèi)IgG2與食蟹猴FcγRIIB的結(jié)合能力高于其與人類(lèi)FcγRIIB的結(jié)合能力;4)人類(lèi)IgG1和IgG3在人類(lèi)或食蟹猴外周血單核細(xì)胞(PBMCs)中介導(dǎo)的ADCC活性相似且較高�,但人類(lèi)IgG4在食蟹猴PBMCs中的活性略高于在人類(lèi)PBMCs中的活性;5)盡管未修飾的人類(lèi)治療性單抗在人類(lèi)和食蟹猴中的效應(yīng)功能譜大致相似���,但Fc修飾可能導(dǎo)致種屬間效應(yīng)特性的差異���,例如去巖藻糖基化的人類(lèi)IgG1與食蟹猴FcγRIIIA結(jié)合時(shí)表現(xiàn)出增強(qiáng)的ADCC活性。
嚙齒類(lèi)動(dòng)物
與人類(lèi)和非人靈長(zhǎng)類(lèi)動(dòng)物相比����,嚙齒類(lèi)動(dòng)物的FcγR在表達(dá)和功能上存在顯著差異����,這增加了小鼠模型使用的復(fù)雜性����。
關(guān)鍵差異包括:小鼠不表達(dá)FcγRIIIB,因此其粒細(xì)胞(中性粒細(xì)胞)缺乏該受體����;小鼠NK細(xì)胞表達(dá)mFcγRIII(更類(lèi)似于人類(lèi)hFcγRIIA),而人類(lèi)NK細(xì)胞表達(dá)hFcγRIIIA(類(lèi)似于小鼠FcγRIV)����;小鼠巨噬細(xì)胞上的FcγRIV是ADCC的關(guān)鍵介導(dǎo)者(人類(lèi)ADCC主要由NK細(xì)胞上的FcγRIIIA介導(dǎo)),實(shí)際上����,小鼠體內(nèi)單核細(xì)胞或巨噬細(xì)胞是負(fù)責(zé)ADCC/ADCP的主要免疫細(xì)胞。小鼠血小板不表達(dá)FcγR(人類(lèi)血小板表達(dá)FcγRIIA)�����。
結(jié)合強(qiáng)度:人類(lèi)IgG與小鼠FcγRs的結(jié)合強(qiáng)度與其同人源同源受體的結(jié)合強(qiáng)度非常相似����,相對(duì)親和力為IgG3>IgG1>IgG4>IgG2����,且FcγRI>>FcγRIV>FcγRIII>FcγRIIB��。
ADCC活性評(píng)估局限:小鼠可能低估人類(lèi)IgG1的ADCC活性���。人類(lèi)IgG1是誘導(dǎo)小鼠效應(yīng)細(xì)胞(通過(guò)小鼠FcγRIII)介導(dǎo)NK細(xì)胞ADCC作用的最強(qiáng)人源同種型,但不如小鼠IgG2a強(qiáng)效���;去巖藻糖基化的人類(lèi)IgG1與小鼠FcγRIII和FcγRIV結(jié)合時(shí)��,比與人類(lèi)hFcγRIIIA結(jié)合時(shí)表現(xiàn)出更強(qiáng)的ADCC活性�����。
其他功能差異:只要小鼠擁有巨噬細(xì)胞����,人類(lèi)IgG1就能在小鼠體內(nèi)介導(dǎo)ADCP��;人類(lèi)IgG2單抗在小鼠和人類(lèi)中激活的效應(yīng)細(xì)胞群不同�����;人類(lèi)IgG4能強(qiáng)效激活小鼠巨噬細(xì)胞(而在人類(lèi)中更惰性);小鼠可能表現(xiàn)出IgG驅(qū)動(dòng)的類(lèi)過(guò)敏反應(yīng)(由血小板活化因子(PAF)和中性粒細(xì)胞介導(dǎo))��,而這在人類(lèi)中并不多見(jiàn)���。
血小板激活風(fēng)險(xiǎn)評(píng)估限制:由于小鼠血小板不表達(dá)FcγR�����,除非使用為此目的構(gòu)建的轉(zhuǎn)基因小鼠�,否則無(wú)法在標(biāo)準(zhǔn)小鼠中評(píng)估血小板激活的潛在風(fēng)險(xiǎn)��。
補(bǔ)體系統(tǒng)比較:已觀察到小鼠介導(dǎo)的CDC活性較弱�����,其原因尚不完全明確�。
FcRn的表達(dá)與功能
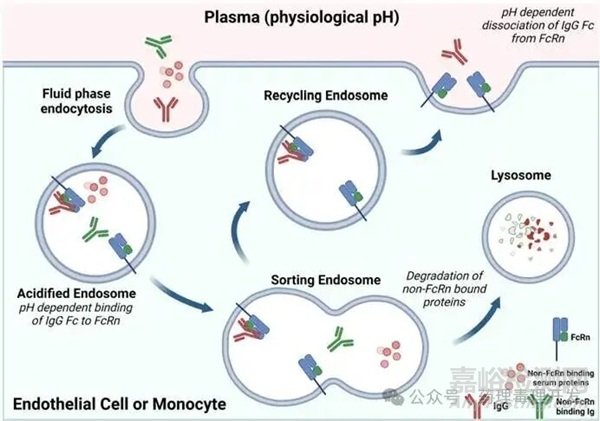
FcRn在免疫和非免疫組織中均有表達(dá),對(duì)IgG的循環(huán)利用和跨細(xì)胞轉(zhuǎn)運(yùn)至關(guān)重要����。FcRn是由MHC-I類(lèi)樣重鏈和β2-微球蛋白輕鏈組成的異二聚體,但該受體不含能進(jìn)行信號(hào)轉(zhuǎn)導(dǎo)或肽呈遞的結(jié)構(gòu)域����,也不與這類(lèi)結(jié)構(gòu)域相關(guān)聯(lián)����。
對(duì)IgG半衰期的影響
FcRn缺陷小鼠的mIgG半衰期從9天縮短至1.4天����,減少了6倍以上,而導(dǎo)入人FcRn轉(zhuǎn)基因可逆轉(zhuǎn)這一效應(yīng)�,這表明FcRn對(duì)IgG半衰期至關(guān)重要。
分布與結(jié)合機(jī)制
分布:FcRn主要存在于內(nèi)體中���,細(xì)胞膜上的表達(dá)量很少(約10%)。
結(jié)合條件:IgG的Fc區(qū)域與FcRn的相互作用發(fā)生在弱酸性pH環(huán)境下(如晚期內(nèi)體中)�,由Fc上的關(guān)鍵組氨酸介導(dǎo)。在內(nèi)體的酸性環(huán)境中��,F(xiàn)cRn與IgG緊密結(jié)合�,使IgG循環(huán)回血液,在生理pH條件下釋放���。
關(guān)鍵作用細(xì)胞與其他功能
半衰期調(diào)控細(xì)胞:血管內(nèi)皮細(xì)胞和骨髓來(lái)源的細(xì)胞(如單核細(xì)胞)是調(diào)控IgG半衰期的關(guān)鍵因素�����。
胎盤(pán)轉(zhuǎn)運(yùn):FcRn能在妊娠期間主動(dòng)將IgG跨胎盤(pán)轉(zhuǎn)運(yùn)�����,使母體IgG傳遞給胎兒�,“新生兒Fc受體”也因此得名�。
肝臟表達(dá):肝臟肝細(xì)胞、庫(kù)普弗細(xì)胞和血竇內(nèi)皮細(xì)胞均已被證實(shí)表達(dá)FcRn�。
種屬間結(jié)合差異與影響
結(jié)合強(qiáng)度:在整個(gè)內(nèi)體pH梯度范圍內(nèi),人類(lèi)IgG與人類(lèi)和小鼠FcRn的結(jié)合強(qiáng)度均高于小鼠IgG�,這對(duì)野生型和人FcRn轉(zhuǎn)基因小鼠的藥理學(xué)和PK評(píng)估有重要影響。
轉(zhuǎn)基因小鼠特點(diǎn):野生型人類(lèi)IgG在常規(guī)小鼠中可被循環(huán)利用����,但當(dāng)進(jìn)行Fc工程改造以延長(zhǎng)血漿半衰期時(shí),其與小鼠FcRn的pH依賴性結(jié)合會(huì)受損����,導(dǎo)致半衰期縮短,而人FcRn轉(zhuǎn)基因小鼠中則無(wú)此情況�;由于人FcRn不結(jié)合小鼠IgG,人FcRn轉(zhuǎn)基因小鼠體內(nèi)的小鼠IgG水平非常低��。因此,在這類(lèi)轉(zhuǎn)基因小鼠模型中評(píng)估IgG單抗制劑時(shí)�,需考慮這一因素——除非在半衰期研究前用靜脈注射免疫球蛋白(IVIG)等混合人IgG處理小鼠,否則不會(huì)存在FcRn結(jié)合的天然競(jìng)爭(zhēng)�。
種間親和力差異:人類(lèi)IgG1與食蟹猴FcRn的親和力比與人FcRn高2倍,與小鼠和大鼠FcRn的親和力比與人FcRn高10倍�����。
FcRn與白蛋白的相互作用
FcRn還可通過(guò)與IgG不重疊的獨(dú)立結(jié)合位點(diǎn)結(jié)合并循環(huán)利用白蛋白�����,從而不僅延長(zhǎng)IgG的血漿半衰期����,也延長(zhǎng)白蛋白的血漿半衰期����。但二者的化學(xué)計(jì)量比不同:一個(gè)FcRn結(jié)合一個(gè)白蛋白����,而兩個(gè)FcRn分子可以“倒置”方式結(jié)合一個(gè)Fc,使Fab段靠近膜����。
人類(lèi)中Fc效應(yīng)功能的不良影響
FcγR和補(bǔ)體的過(guò)度激活可能給含F(xiàn)c的治療藥物帶來(lái)潛在安全風(fēng)險(xiǎn)�����。
不必要的FcγR陽(yáng)性細(xì)胞激活會(huì)驅(qū)動(dòng)ADCC����、ADCP及C1q/CDC介導(dǎo)的靶細(xì)胞殺傷,可能導(dǎo)致免疫系統(tǒng)異常激活和細(xì)胞因子釋放(包括腫瘤溶解綜合征及其他后遺癥)�����,OKT3�����、利妥昔單抗�、奧濱尤妥珠單抗和阿侖單抗的應(yīng)用中均觀察到此類(lèi)現(xiàn)象。
含F(xiàn)cγR的免疫細(xì)胞不必要的交聯(lián)和激活���,可能導(dǎo)致與靶標(biāo)生物學(xué)特性相關(guān)的激活依賴性凋亡�����、邊緣池化��、增殖�����、抑制或內(nèi)化�。
循環(huán)中多價(jià)可溶性分子的交聯(lián)可能導(dǎo)致可溶性免疫復(fù)合物形成,引發(fā)炎癥反應(yīng)����。
與血小板上的FcγRIIA結(jié)合可能促進(jìn)血小板激活,并可能引發(fā)血栓事件���,在動(dòng)物和人類(lèi)中使用抗CD40L單抗時(shí)已觀察到這種情況���。
補(bǔ)體激活可產(chǎn)生C3a和C5a(過(guò)敏毒素),誘導(dǎo)表達(dá)C3aR/C5aR的肥大細(xì)胞�、嗜堿性粒細(xì)胞和內(nèi)皮細(xì)胞異常激活。